

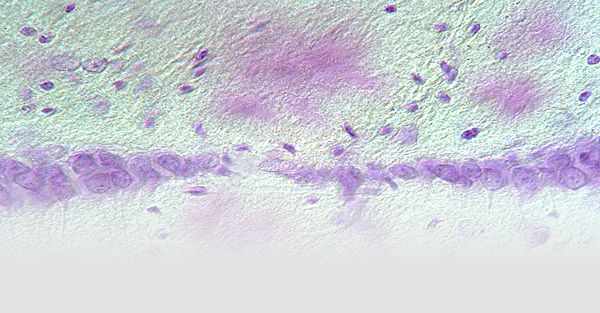
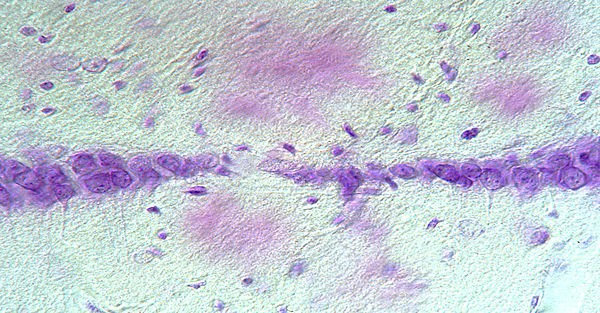
Cerebral proteopathy is a unifying term for neurodegenerative diseases in which aggregated proteins are abnormally deposited in the brain. The hallmark proteopathy is Alzheimer’s disease (AD). In AD a key early event is the misfolding and aggregation of the amyloid-β peptide (Aβ). What the initial trigger is for the misfolding and how the aggregation of disease-specific proteins leads to neurodegeneration is largely unknown.
1. To understand how Aβ aggregation originates, spreads, and leads to neuronal dysfunction and dementia.
2. To study non-Aβ proteopathies in order to determine their similarities and differences relative to the Aβ-type.
To this end, we are using organotypic slice cultures and a variety of genetically engineered mouse models of cerebral β-amyloidoses, tauopathies and α-synucleinopathies. To foster the translational and therapeutic aspects of our work, the results obtained from our mouse models are analyzed in comparison to those in the respective human patient samples.
One seminal finding from our group was the observation that dilute extracts of Aβ-containing material, either from the brains of AD patients or from aged Aβ precursor protein- (APP) transgenic mice, are able to induce cerebral β-amyloidosis and associated lesions in the brains of young APP-transgenic mice (Meyer-Lühmann et al., Science 2006; Eisele et al., Science 2010). When Aβ is biochemically inactivated or removed from the brain samples, the extracts lose the ability to induce Aβ deposition, showing that Aβ itself is necessary for the amyloid induction. Surprisingly, however, synthetic Aβ is much less efficient in inducing the amyloid lesions, suggesting that the ability of Aβ to induce cerebral β-amyloidosis requires the Aβ molecules to acquire certain structural characteristics or co-factors that are generated in the living brain (Aβ seeds). While the self-propagating nature of Aβ seeds bears important similarities to that of prions, there is currently no evidence that AD is transmissible in the same sense as are prion diseases (Jucker and Walker, Nature Neurosci 2018).
The prion-paradigm, however, implies that the conformational characteristics of Aβ seeds govern the type and toxicity of Aβ deposition. Indeed, we have found evidence that the various clinical features of AD may result from different molecular Aβ conformations (Rasmussen et al., PNAS 2017). Moreover, the self-propagating nature of Aβ aggregates raises the possibility that Aβ seeds could serve as a very early diagnostic biomarker for cerebral β-amyloidosis and maybe AD. This idea is further supported by previous findings that the most potent Aβ seeds are small and soluble and may thus occur in bodily fluids.
Similar to Aβ seeds, α-synuclein seeds are not only very active but also durable (Schweighauser et al., Acta Neuropathol 2015). The discovery that proteopathic seeds can persist in the brain for months at levels below routine detection indicates that the initial changes that drive AD pathogenesis may occur even earlier than previously thought (Ye et al., Nat Neurosci 2015; Ye et al., EMBO Rep 2017; Uhlmann et al., Nature Neurosci 2020). From a therapeutic standpoint, these findings indicate that the most effective treatments for AD should target the Aβ cascade well before the characteristic deposits of Aβ become detectable by currently available methods (Schelle et al., Ann Neurol 2019; Uhlmann et al., Nature Neurosci 2020).

+49 (0)7071-
29-81945

+49 (0) 7071-
29-86863

+49 (0)7071-
29-81946

+49 (0) 7071-
29-86851

+49 (0)7071-
29-81947

+49 (0)7071-
29-81924

+49 (0) 7071-
29 -81945
Full list of publications
Selected publications
Jucker M, Walker LC (2024) Evidence for the iatrogenic transmission of Alzheimer's disease. Nature Medicine 30(2):344-345 (Abstract)
Jucker M, Walker LC (2023) Alzheimer's disease: From immunotherapy to immunoprevention. Cell 186(20):4260-4270 (Abstract)
Rother C, Uhlmann RE, Müller SA, Schelle J, Skodras A, Obermüller U, Häsler LM, Lambert M, Baumann F, Xu Y, Bergmann C, Salvadori G, Loos M, Brzak I, Shimshek D, Neumann U, Dominantly Inherited Alzheimer Network, Walker LC, Schulz SA, Chhatwal JP, Kaeser SA, Lichtenthaler SF, Staufenbiel M, Jucker M (2022) Experimental evidence for temporal uncoupling of brain Aβ deposition and neurodegenerative sequelae. Nat Commun 13:7333 (Abstract)
Uhlmann RE, Rother C, Rasmussen J, Schelle J, Bergmann C, Ullrich Gavilanes EM, Fritschi SK, Buehler A, Baumann F, Skodras A, Al-Shaana R, Beschorner N, Ye L, Kaeser SA, Obermüller U, Christensen S, Kartberg F, Stavenhagen JB, Rahfeld JU, Cynis H, Qian F, Weinreb PH, Bussiere T, Walker LC, Staufenbiel M,Jucker M (2020) Acute targeting of pre-amyloid seeds in transgenic mice reduces Alzheimer-like pathology later in life. Nat Neurosci 23(12):1580-8 (Abstract)
Schelle J*, Wegenast-Braun BM*, Fritschi SK, Kaeser SA, Jährling N, Eicke D, Skodras A, Beschorner N, Obermueller U, Häsler LM, Wolfer DP, Mueggler T, Shimshek DR, Neumann U, Dodt H-U, Staufenbiel M, Jucker M (2019) Early Aβ reduction prevents progression of cerebral amyloid angiopathy. Ann Neurology 86(4):561-71 (Abstract)
Jucker M, Walker LC (2018) Propagation and spread of pathogenic protein assemblies in neurodegenerative diseases. Nat Neurosci 21:1341-1349 (Abstract)
Rasmussen J*, Mahler J*, Beschorner N*, Kaeser SA, Häsler LM, Baumann F, Nystrom S, Portelius E, Blennow K, Lashley T, Fox NC, Sepulveda-Falla D, Glatzel M, Oblak AL, Ghetti B, Nilsson KPR, Hammarstrom P, Staufenbiel M, Walker LC, Jucker M (2017) Amyloid polymorphisms constitute distinct clouds of conformational variants in different etiological subtypes of Alzheimer's disease. Proc Natl Acad Sci USA 114:13018-23 (Abstract)
Ye L*, Rasmussen J*, Kaeser SA, Marzesco AM, Obermuller U, Mahler J, Schelle J, Odenthal J, Kruger C, Fritschi SK, Walker LC, Staufenbiel M, Baumann F, Jucker M (2017) Abeta seeding potency peaks in the early stages of cerebral beta-amyloidosis. Embo Reports 18:1536-44 (Abstract)
Ye L*, Fritschi SK*, Schelle J, Obermüller U, Degenhardt K, Kaeser SA, Eisele YS, Walker LC, Baumann F, Staufenbiel M, Jucker M (2015) Persistence of Aβ seeds in APP null mouse brain. Nat Neurosci 18:1559-61 (Abstract)
Schweighauser M, Bacioglu M, Fritschi SK, Shimshek DR, Kahle PJ, Eisele YS, Jucker M (2014) Formaldehyde-fixed brain tissue from spontaneously ill alpha-synuclein transgenic mice induces fatal alpha-synucleinopathy in transgenic hosts. Acta Neuropathol 129:157-9 (Abstract)
Fritschi SK, Langer F, Kaeser S, Maia LF, Portelius E, Pinotsi D, Kaminski CF, Winkler DT, Maetzler W, Keyvani K, Spitzer P, Wiltfang J, Kaminski Schierle GS, Zetterberg H, Staufenbiel M, Jucker M (2014) Highly potent soluble amyloid-β seeds in human Alzheimer brain but not cerebrospinal fluid. Brain 137:2909-15 (Abstract)
Jucker M, Walker LC (2013) Self-propagation of pathogenic protein aggregates in neurodegenerative diseases. Nature 501:45-51 (Abstract)
Eisenberg D, Jucker M (2012) The amyloid state of proteins in human diseases. Cell 148:1188-203 (Abstract)

Hertie Center of Neurology
Hertie Institute for Clinical Brain Research
Department Cellular Neurology
Otfried-Müller-Straße 27
D-72076 Tübingen
Gisela Rose
Phone: +49 (0)7071 29-81947
Fax: +49 (0)7071 29-4521
gisela.roseuni-tuebingen.de
Dr. Jörg Odenthal
Phone: +49 (0)7071 29-86862
Fax: +49 (0)7071 29-4757
joerg.odenthaluni-tuebingen.de




